Globala klimatförändringar
och korallblekning
 |
Många skulle säkert klassa
störningar relaterade till globala klimatförändringar
som ett resultat av mänskliga aktiviteter, och
med tanke på den ökande växthuseffekten är det
troligtvis också korrekt att göra det. Jag har
dock valt att behandla hot relaterade till klimatförändringar
i en egen del av hemsidan, mycket beroende på
att det finns så mycket att skriva om det.
Korallrev är mycket känsliga
mot förändringar i klimatet, och korallblekning,
som är den allvarligaste och mest utbredda effekten
av den globala uppvärmningen, kan i extremfall
döda koraller och omvandla korallrev till andra,
mer lågproduktiva och artfattiga ekosystem. Korallblekning
är dock något som inte bara inträffar vid temperaturförändringar.
Exempelvis har ljusintensitetens inverkan fått
en allt större uppmärksamhet på sistone, och sediment,
sötvatten och toxiska ämnen kan alla inducera
korallblekning, även om det vanligtvis bara sker
lokalt. Det som sker vid korallblekning är att
korallernas zooxantheller stöts ut och/eller förlorar
sitt fotosyntetiserande pigment med försvagade
koraller som resultat.
Korallblekning började först
studeras under 1970-talet då man undersökte hur
toleranta koraller är mot höga vattentemperaturer,
men det var inte förrän korallblekning blev ett
globalt fenomen under 1980- och 1990-talet (på
grund av den globala uppvärmningen och ENSO) som
man förstod vilken effekt det har på koraller
och korallrev.
En
introduktion: Korallblekning
Vid korallblekning förlorar korallerna sin färg
och det underliggande vita kalkskelett blir synligt.
Mer exakt är det korallernas endosymbionter, det
vill säga zooxanthellerna, som stöts ut och/eller
förlorar sitt fotosyntetiserande pigment. Av de
tusentals zooxantheller som återfinns per kubikcentimeter
korallpolyp förlorar korallerna vanligtvis 60-90
procent av dessa vid korallblekning samtidigt
som de enskilda zooxanthellerna kan förlora 50-80
procent av sina fotosyntetiserande pigment. Då
en så stor mängd zooxantheller försvinner och/eller
förlorar sitt fotosyntetiserande pigment förlorar
korallerna sin färg och det vita kalkskelettet
lyser igenom de genomskinliga korallpolyperna.
Koraller som även har andra, fluorescerande pigment
som inte är associerade med zooxanthellernas fotosyntetiserande
pigment behöver dock inte förlora all färg. I
samband med korallblekning kan dessa pigment ibland
bli mer intensiva i sitt framträdande, och skarpa
pastellfärger blandas då upp med de annars blekta
korallerna. De fluorescerande proteiner som bygger
upp dessa pigment kan emellertid också försvinna
eller minska i antal, vilket resulterar i näst
intill fullständig korallblekning (alla zooxantheller
försvinner som sagt inte). Pigmenten assisterar
zooxanthellernas fotosyntes genom att förbättra
ljusupptaget, men försvinner zooxanthellerna förlorar
de sin roll, och eftersom det är kostsamt för
värden att producera dem försvinner även de efter
en viss tid. Blekning har också påträffats hos
andra organismer som lever i symbios med zooxantheller,
exempelvis mjukkoraller och jättemusslor.
Korallblekning kan induceras
av flera olika typer av stress med temperaturförändringar
som den vanligaste faktorn. Koraller är anpassade
till att leva i en förhållandevis smal temperaturmarginal,
och både för låga och för höga ytvattentemperaturer
kan resultera i korallblekning. Intensiva uppvällningsperioder
kan exempelvis resultera i att vattentemperaturen
plötsligt sjunker med 3-5°C, och det kan få korallerna
att blekna inom bara 5-10 dagar. Korallblekning
i samband med förhöjda vattentemperaturer är dock
mer utbrett, och en så liten temperaturhöjning
som 1-2°C över korallernas temperaturmarginal
(tröskelnivå) under 5-10 veckor på sommarhalvåret
brukar leda till korallblekning. Dessa förhöjda
vattentemperaturer behöver dock inte alltid vara
ett resultat av vädret utan också termisk förorening
från kraftverk kan framkalla korallblekning. Solinstrålning
i form av vanligt synligt ljus och UV-ljus har
också visat sig kunna inducera korallblekning,
speciellt i kombination med förhöjda vattentemperaturer
(se Korallblekning
relaterad till förhöjda vattentemperaturer).
Om koraller exponeras för luft under perioder
med stark solinstrålning och värme vid exempelvis
lågt tidvatten kan de också blekna, och i ett
fåtal fall har man kunnat koppla korallblekning
till sedimentation. Snabb utspädning av havsvatten
(sänkt salthalt) i samband med regnoväder och
sötvattenavrinning från land kan också inducera
korallblekning i små kustnära områden, men vanligast
är dock att en sänkt salthalt (< 23 promille)
inte bara resulterar i en förlust av zooxantheller,
utan i att hela vävnaden fälls av, vilket exponerar
korallernas underliggande kalkskelett. Större
utsläpp av toxiska ämnen som exempelvis olja är
ytterligare en typ av stress som kan resultera
i korallblekning, även om det i så fall endast
handlar om en mycket lokal effekt. Ökade halter
av oorganiska näringsämnen (eutrofiering) inducerar
däremot inte korallblekning utan förhöjer istället
koncentrationerna av zooxantheller i korallernas
vävnad. Däremot kan föroreningar av det här slaget
försämra korallernas resistens och förvärra effekterna
av korallblekning. Även bakteriella infektioner
har visat sig resultera i korallblekning hos vissa
arter (se Korallsjukdomar),
och koraller som exponeras för natriumcyanid (se
Destruktiva
fiskemetoder) kan drabbas av allt från lindrig
till fullständig korallblekning.
Blekta koraller växer mycket
mer långsamt än friska eftersom det energitillskott
de vanligtvis får från sina endosymbionter försvinner,
och de kan bli mer mottagliga för sjukdomar och
andra störningar. Vid mild korallblekning överlever
emellertid korallerna som oftast och de kan börja
återuppbygga sina populationer av zooxantheller
inne i korallpolyperna. Det sker antingen genom
upptag av nya zooxantheller från vattnet och/eller
genom reproduktion av de kvarvarande zooxanthellerna
inne i korallpolypernas vävnad. Eftersom de fluorescerande
pigmenten då återigen behövs reproduceras även
dessa. Återhämtningen från korallblekning är dock
beroende av ett flertal olika faktorer, exempelvis
storleken på korallernas energiförråd och deras
förmåga att fånga egen föda under tiden zooxanthellerna
återuppbygger sina populationer.
Vid extrema förhållanden kan
emellertid blekta koraller dö, men i de fall där
mortliteten endast är partiell kan kvarvarande
korallfragment återhämta sig och fortsätta tillväxa,
även om det tar 1-2 år innan de har återfått normal
reproduktions- och tillväxtkapacitet. Skulle ett
helt korallrev drabbas av korallblekning med massmortalitet
bland korallerna som resultat kan det emellertid
ta flera decennier för korallrevet att återhämta
sig helt. Vid en mindre omfattande mortalitet
(flertalet koraller överlever helt eller partiellt)
kan en signifikant återhämtning dock ske redan
inom 2-3 år.
Korallblekning
relaterad till förhöjda vattentemperaturer
Vetenskapliga studier har nu bekräftat att jordens
yttemperatur har stigit med 0,6°C under de senaste
100 åren, och det är den snabbaste ökningen på
1 000 år. Mycket tyder dessutom på att dagens
förhöjda luft- och vattentemperaturer är ett resultat
av mänskliga aktiviteter som exempelvis skogsskövling
och förbränning av fossila bränslen, aktiviteter
som påverkar tillförseln av växthusgaser till
atmosfären och förstärker växthuseffekten.
Det är troligt att väderfenomenet
ENSO (El Niño-Southern Oscillation), som förekommer
med några års mellanrum, har blivit kraftigare
på grund av den globala uppvärmningen. Som namnet
antyder är fenomenet uppdelat i två separata delar;
"Southern Oscillation" och "El Niño". "Southern
Oscillation"-delen innebär att den atmosfäriska
tryckskillnaden mellan Stilla Havet och Indiska
Oceanen kastas om, vilket resulterar i att vindmönstret
och nederbörden förändras, och att den blöta sommarmonsunen
i södra Asien uteblir. "El Niño"-delen innebär
att passadvindarna längs med Sydamerikas västkust
avstannar, vilket resulterar i att uppvällningen
upphör helt, och att ytvattentemperaturen stiger
kraftigt. Det var först under 1950-talet som det
blev uppenbart att "El Niño" och "Southern Oscillation"
egentligen var två sidor av samma mynt, och att
de tillsammans är del av ett komplex samspel mellan
havet och atmosfären som sammankopplar hela planeten.
Därför använder man sig ofta av förkortningen
ENSO för att symbolisera att "El Niño" och "Southern
Oscillation" tillsammans skapar de globala klimatförändringar
man kan se med några års mellanrum.
Vanligtvis förekommer ENSO i
12-18 månader långa perioder och har som sagt
en global inverkan. Västra Stilla Havet drabbas
av torka medan östra Stilla Havet drabbas av kraftig
nederbörd och översvämningar, och under 1997-98
års ENSO, som kan ha varit den kraftigaste i historien,
drabbades även centrala Europa av översvämningar.
Även havet påverkas, och det är speciellt under
ENSO-år som så kallade massblekningar inträffar
på grund av att ytvattentemperaturen stiger globalt.
Vid en massblekning drabbas stora delar av ett
korallrev till hela system av korallrev av korallblekning,
och antalet massblekningar har ökat stadigt sedan
de första registrerades 1979. De hittills mest
omfattande massblekningarna inträffade i samband
med 1997-98 års ENSO och beskrivs i mer detalj
nedan.
Det är möjligt att den globala
uppvärmningen också påverkar andra atmosfäriska
och oceaniska mönster. En förändrad styrka och
riktning hos havsströmmarna skulle exempelvis
kunna påverka de lokala ytvattentemperaturerna,
och förändringar i den atmosfäriska cirkulationen
som ett resultat av ett förändrat klimat kan påverka
uppvällningen och frekvensen och intensiteten
hos regionala väderextremer. Alla dessa faktorer
kan potentiellt öka antalet massblekningar och
göra dem mer intensiva, men det finns fortfarande
en stor osäkerhet kring dessa förändringars lokalisering,
magnitud och riktning.
Massblekningen
1997-98
Under 1997-98 års ENSO drabbades korallrev i hela
världen av massblekningar och uppskattningsvis
16 procent av alla korallrev skadas allvarligt
(se Karta 3 i Kartbanken).
Värst drabbades korallreven i centrala Indiska
Oceanen där uppemot 90 procent av korallerna på
flera tusen kvadratkilometer korallrev dog, inklusive
praktiskt taget alla korallrev på Maldiverna,
Chagosarkipelagen och Seychellerna. Även andra
korallrev, exempelvis utanför Tanzania och Bahrain,
drabbades av svår korallblekning med en massmortalitet
bland korallerna som ofta nådde 95 procent. På
korallrev utanför exempelvis Thailand, Vietnam
och Kenya dog 50-70 procent av korallerna, men
här kunde också mer massiva och motståndskraftiga
korallarter återhämta sig. Delar av Stora Barriärrevet,
Indonesien, Filippinerna och Karibien drabbades
också av korallblekning under massblekningen 1997-98
med en mortalitet bland korallerna på runt 20-50
procent. I andra områden som Röda Havet, södra
Indiska Oceanen och centrala Stilla Havet kunde
man däremot inte observera någon betydande korallblekning
under dessa ENSO-år, och det samma gäller för
stora delar av Stora Barriärrevet. I vissa opåverkade
områden hade emellertid massblekningar förekommit
tidigare.
Massblekningen pågick från mitten
av 1997 till slutet av 1998 och sammanträffade
som sagt med en extremt kraftig ENSO som sedan
snabbt övergick till en kraftig La Niña, ett väderfenomen
som i stort sett medför väderförhållanden motsatta
El Niños. Det här var troligtvis den varmaste
perioden som moderna koraller har fått utstå,
och den drabbade främst koraller i vatten grundare
än 15 meter. Speciellt förgrenade och snabbväxande
koraller som Acropora påverkades mest,
men även massiva och långsamt växande koraller
drabbades. Många av de senare kunde dock återhämta
sig inom 1-2 månader.
De värst drabbade korallreven
i Indiska Oceanen kommer att behöva år eller decennier
på sig för att kunna återhämta sig helt, men korallrev
i exempelvis Seychellerna, som efter massblekningen
inte täcktes av levande koraller till mer än 5
procent, har uppvisat en anmärkningsvärd återhämtning,
och andelen levande koraller var 2004 uppe i 20-30
procent. Korallrev i områden som Chagosarkipelagen
och Sri Lanka har emellertid inte uppvisat en
lika positiv återhämtning, och även utanför Indiska
Oceanen ser man en tydlig variation i återhämtningen
hos drabbade korallrev. Återhämtningen sker genom
kolonisering av nya planula-larver och ny tillväxt
hos koraller som tidigare ansågs vara döda, men
för att en fullständig återhämtning skall kunna
äga rum krävs det att massblekningar med en liknande
intensitet inte upprepas och att drabbade korallrev
samtidigt inte stressas av mänskliga aktiviteter,
och sammantaget är ett korallrevs återhämtning
från korallblekning och massmortalitet en komplex
process. Först och främst är den beroende av att
lämpliga larver framgångsrikt settlar, överlever
och tillväxer, och unga koraller får utstå både
predation och konkurrens från andra organismer,
inklusive andra koraller. Dessutom måste det finnas
ett tillflöde av planula-larver från andra korallrev
uppströms (ny forskning har dock visat att larvtillförseln
ofta kan vara mer lokal), och på dessa korallrev
måste möjligheterna till att reproducera sig i
sin tur vara goda. Tillsammans resulterar dessa
osäkerheter i att ett korallrevs återhämtning
till den diversitet och rikedom av koraller som
återfanns innan blekningen troligtvis blir en
uppgift som sträcker sig över flera decennier.
Trots en viss återhämtning från
massblekningen 1997-98 är man dock orolig för
att massmortaliteter av den här magnituden kan
leda till lokal utrotning av vissa arter och att
det i sin tur resulterar i minskad biodiversitet
och förändringar inom ekosystemet. Och man har
all anledning att vara orolig; jordens yttemperatur
förväntas stiga med mellan 0,6 och 4°C under de
närmaste 100 åren, samtidigt som temperaturen
i tropiska hav för tillfället stiger med uppskattningsvis
1-2°C per sekel, och om 30-50 år kommer troligtvis
massblekningar liknandes den 1997-98 att inträffa
årligen, oberoende av ENSO.
De
biokemiska processerna bakom blekningen
Förhöjda vattentemperaturer kan ensamt förklara
flera fall av massblekningar, men det finns också
en variation associerad med massblekningar som
inte kan förklaras av förändrade ytvattentemperaturer,
exempelvis att de ytligaste delarna av en korall
oftast bleks först och med störst intensitet,
och att koraller som står bredvid varandra inte
alltid har samma benägenhet att blekas. Därför
trodde man att andra faktorer spelade in, och
de som förespråkades mest var korallers och zooxanthellers
genotyp (genetiska egenskaper) och ljusintensiteten.
Att olika koraller reagerar olika på förändringar
i ytvattentemperaturen är välkänt och beror på
att vissa koraller helt enkelt är mer värmetåliga
än andra (se Resistens),
men ljusintensitetens inverkan på korallblekning
fick också en stor uppmärksamhet. Vissa fall av
korallblekning har föregåtts av perioder med klart
och lugnt vatten som underlättar solinstrålningen
av vanligt synligt ljus och UV-ljus, och i experiment
med bland annat koraller har blekning kunnat induceras
under förhöjda nivåer av UV-ljus. Stark solinstrålning
under normala temperaturförhållanden kan dock
inte ensamt skapa massblekningar, så en kombination
av stark solinstrålning och förhöjd ytvattentemperatur
skulle därför vara en rimlig orsak till massblekningar.
Senare studier har också visat att en förhöjd
vattentemperatur och solinstrålning tillsammans
skadar zooxanthellernas fotosyntes, och det är
det som resulterar i den vanligaste formen av
korallblekning.
Fotosyntesen äger rum i zooxanthellernas
kloroplatser som är uppbyggda av ett yttre och
ett inre biologiskt membran. Innanför det inre
membranet återfinns så kallade tylakoidmembran
där proteinerna som sköter fotosyntesens ljusreaktioner
är lokaliserade, och av dessa proteiner är kanske
Fotosystem I och Fotosystem II de mest välkända.
Fotosystem II absorberar först ljus (fotoner)
med hjälp av antennpigment, och exciterar därefter
elektroner (från vattenmolekyler) som får transporteras
genom en elektrontransportkedja till Fotosystem
I där de exciteras igen. Därpå kommer de exciterade
elektronerna att ingå i reducerande molekyler
som har en viktig roll i fotosyntesens mörka reaktioner
(där beståndsdelarna till mer avancerade organiska
föreningar tillverkas). Även ATP (cellens "energivaluta")
och syre produceras i fotosyntesens ljusreaktioner.
Man har visat att de exciterade
elektronerna inte längre når fram till fotosyntesens
mörka reaktioner vid förhöjda temperaturer, och
att de istället reagerar med syre och producerar
aktiva syreradikaler. De aktiva syreradikalerna
börjar sedan denaturera de proteiner som bygger
upp komponenterna i zooxanthellernas fotosyntes,
och ljusreaktionerna förstörs. I princip är det
alltså det ljus, som de flesta koraller är beroende
av för att kunna överleva, som gör att korallerna
bleknar vid förhöjda vattentemperaturer eftersom
det fortsätter att excitera elektroner även efter
det att de mörka reaktionerna inte längre kan
nås. Korallblekning är därmed relaterat till fenomenet
fotoinhibering som förekommer hos fotosyntetiserande
organismer i både vatten och på land. Vid fotoinhibering
absorberas mer ljusenergi än vad fotosyntesen
kan ta hand om, och överskottsenergin börjar då
producera potentiellt farliga produkter, exempelvis
aktiva syreradikaler, som reducerar fotosynteskapaciteten.
Zooxantheller har ett enzymatiskt skydd som detoxifierar
dessa syreradikaler, men trots detta verkar alltså
korallblekning uppstå som ett resultat av en sänkt
känslighet för fotoinhibering hos korallernas
endosymbionter. De fluorescerande pigment som
återfinns i korallpolypernas vävnad skulle dock
kunna fungera som ett kompletterande skydd mot
korallblekning genom att filtrera bort UV-ljus
och överflödigt synligt ljus (se Resistens
och Kan
inte korallerna anpassa sig?).
Sammanfattningsvis resulterar
alltså en förhöjd vattentemperatur i att zooxanthellernas
fotosyntes havererar, och att skadliga aktiva
syreradikaler skapas då ljusinstrålningen fortsätter.
De aktiva syreradikalerna bryter därefter ner
fotosyntesen, och det är då zooxanthellerna förlorar
sitt fotosyntetiserande pigment. Troligtvis stöts
zooxanthellerna ut ur korallerna i samband med
att de celler de lever i förlorar sin vidhäftning
till andra celler och drivs ut, något som inträffar
vid både för höga och låga temperaturer. Zooxantheller
kan stötas ut i en förhållandevis kontrollerad
process, men utsätts korallerna för stress kan
alltså korallpolyperna i värsta fall förlora sina
egna celler. Det finns dock en annan hypotes som
menar att utstötningen av zooxantheller kan vara
ett adaptivt beteende från korallernas sida (se
Kan inte
korallerna anpassa sig?), och det har också
föreslagits att korallerna stöter ut sina zooxantheller
som ett resultat av att de inte tål de aktiva
syreradikaler som produceras i samband med fotoinhiberingen;
korallpolypernas cellulära processer kan skadas
av dessa toxiska substanser.
Resistens
Olika koraller är som sagt olika mottagliga för
korallblekning. Vissa massiva arter från släktet
Porites är exempelvis förhållandevis resistenta
mot temperaturstress, och om de skulle blekas
tenderar de att återhämta sig utan någon direkt
mortaliteten. De förgrenade Acropora-korallerna
är dock mer känsliga mot en höjning av ytvattentemperaturen,
och mortaliteten kan i vissa fall ligga på uppemot
95 procent. Koraller som är mer resistenta har
också en tendens att karaktäriseras av solida,
massiva kalkskelett med tjock vävnad och långsam
tillväxthastighet, och här ingår inte Acropora
eller andra släkten med förgrenat kalkskelett.
Varför olika korallarter är
olika känsliga för temperaturstress beror på vilken
typ av zooxanthell de lever i symbios med, ljusmiljön
inne i korallernas vävnad (exempelvis kan en tjock
vävnad skugga zooxanthellerna och öka resistensen)
och externa, fysiska faktorer. Under början av
2000-talet har forskarvärldens fokus dock legat
på diversiteten av zooxantheller. Totalt finns
det 8 olika typer (klader; A, B, C, D, E, F, G
och H) av zooxantheller, och medan vissa lever
i symbios med koraller, är andra inte associerade
med koraller. Typ A förekommer i både stenkoraller
och havsanemoner och anses vara förhållandevis
motståndskraftig mot perioder av korallblekning.
Detsamma gäller för typ B, som bland annat återfinns
i flera släkten av stenkoraller i Atlanten. Typ
C, som är mindre resistent, har påträffats i flera
havsområden, men vanligtvis associeras den med
revbyggande koraller i tropiska Stilla Havet.
I Stilla Havet har också korallblekning ofta noterats
i koraller som lever i symbios med zooxantheller
av typ C. Till skillnad från typ C är typ D förhållandevis
motståndskraftig mot korallblekning, och den återfinns
ofta i områden som nyligen har drabbats av kraftig
korallblekning (se "Adaptive
Bleaching Hypothesis" och acklimatisering).
Av typerna A, B, C och D är också typ D den mest
värmeresistenta typen. Av de återstående typerna
verkar typ E däremot inte förekomma i koraller,
medan typ F återfinns i vissa korallarter. Typ
G har bara nyligen påträffats (år 2005) och återfinns
i mjukkoraller, stenkoraller och stora havsanemoner.
Men varför är vissa zooxantheller
mer motståndskraftiga än andra mot förhöjda vattentemperaturer
och/eller ljusintensiteter? Först och främst kan
vissa zooxantheller skydda sig mot UV-ljus och
vanligt synligt ljus. Exempelvis skyddar xantofyller,
en typ av karotenoida pigment som påträffas hos
både zooxantheller, alger och högre växter, Fotosystem
II mot synligt (fotosyntetiskt aktivt) ljus genom
att ta bort blått ljus, som har visat sig intensifiera
temperaturinducerad korallblekning. Det innebär
att zooxantheller som har förmågan att producera
xantofyller tål högre ljusintensiteter utan att
deras fotosyntes skadas. Liksom synligt ljus kan
som sagt också UV-ljus inducera korallblekning
i samband med förhöjda ytvattentemperaturer, men
även här har vissa zooxantheller ett skydd. Detta
skydd består av en typ av aminosyror som förkortas
MAA (mycosporine-like amino acids), och det skulle
då ge de koraller som har dessa zooxantheller
en klar konkurrensfördel i grunda vatten där solinstrålningen
är stor. Ett ytterligare skydd skulle kunna vara
själva tylakoidmembranets kemiska komposition,
medan mörkpigmenterade koraller, som potentiellt
kan ta upp mer värme och bli varmare än det omgivande
vattnet, kan göra sina zooxantheller mer mottagliga
för temperaturstress, och, följaktligen, sig själva
mer mottagliga för korallblekning.
Fluorescerande pigment kan möjligtvis
också tillhandahålla koraller med en viss resistens
(se Kan
inte korallerna anpassa sig?), men det finns
också mer fysiska skydd, exempelvis skuggning
från moln och öar, eller från en hög turbiditet
(partikelhalt) i vattnet. Korallarter som har
anpassat sig till turbida miljöer är också mer
beroende av att fånga egen föda, och det är möjligt
att de då inte svälter lika mycket som andra arter
gör i samband med korallblekning. Områden med
varierande ljusintensiteter, salthalter, temperaturer
och halter av oorganiska näringsämnen (exempelvis
uppvällningsområden och områden med god vattencirkulation)
kan fungera som ytterligare skydd mot kraftig
korallblekning, och i mindre skala kan även variationer
i mikromiljön påverka känsligheten för värmestress.
Även regionala och lokala skillnader i vädret
kan skapa förutsättningar som påverkar mottagligheten
för korallblekning.
Konsekvenserna
av korallblekning
Sedan 1979, då de första registrerade massblekningarna
inträffade, har man kunnat se vilka effekter korallblekning
har på koraller och korallrev. Nedan följer en
beskrivning av de möjliga effekter som eventuella
framtida klimatförändringar kommer att ha på koraller
och korallrev.
Ökad
dödlighet bland korallerna
En av de mest direkta effekterna som korallblekning
har på koraller och korallrev är att korallerna
tenderar att dö snabbare. Dödligheten kan dock
variera från korallrev till korallrev. På vissa
korallrev kan den exempelvis ligga på närmare
noll procent vid mild korallblekning, medan andra
korallrev kan drabbas av en massmortalitet på
nära 100 procent i samband med massblekningar.
Detta inträffade exempelvis på vissa korallrev
i östra Stilla Havet och Indonesien under 1982-83
års ENSO. Dödligheten bland korallerna beror på
hur länge de utsätts för förhöjda vattentemperaturer
och hur högt temperaturerna når. Därför är det
otvivelaktigt att en framtida global uppvärmning
kommer att leda till längre och intensivare massblekningar,
och att mortaliteten bland korallerna kommer att
öka så pass mycket under de närmaste decennierna
att den till slut kommer att nå nästan fullständig
dödlighet.
Eftersom förekomsten av massblekningar
ökar är det också troligt att dödligheten bland
korallerna för med sig andra problem. Massblekningar
inträffar idag med 3-4 års mellanrum, och det
kan få effekter på den framtida rikedomen av koraller
eftersom de flesta koraller då hinner dö innan
de har hunnit bli könsmogna och kunnat reproducera
sig. Exempelvis tar de förgrenade Acropora-korallerna
uppskattningsvis 4-5 år på sig för att bli könsmogna,
och om massblekningarna blir allt vanligare i
framtiden skulle alltså dessa koraller hinna dö
innan de hunnit reproducera sig. Massiva och mer
långsamt växande koraller som blir könsmogna senare
skulle då drabbas ännu hårdare, och bara de korallarter
som blir könsmogna tidigt under sin livscykel
skulle kunna reproducera sig. Massiva koraller
(exempelvis släktet Porites) är dock mer
motståndskraftiga mot korallblekning, något som
gör det svårt att förutse den framtida artsammansättningen
av koraller på korallrev som har drabbats av massblekningar.
Man skall emellertid inte tro
att dagens korallarter kommer att utrotas av de
förhöjda vattentemperaturerna eftersom koraller
har överlevt långt större klimatförändringar under
de miljoner år de har existerat på jorden.
Försämrad
reproduktion bland korallerna
Förutom att massblekningar kan få koraller att
dö innan de haft möjligheten att reproducera sig
har man också kommit fram till att den reproduktiva
kapaciteten hos blekta koraller minskar. I en
studie som hade för avsikt att undersöka hur korallernas
reproduktiva kapacitetet påverkades av korallblekning
såg man tydliga resultat; de flesta av de 200
koraller från grunda vatten utanför Heron Island,
Australien, som undersöktes efter massblekningen
1997-98 hade fått sin reproduktiva aktivitet reducerad
av blekningen. Blekta koraller från flera viktiga
arter saknade helt äggceller, även om de skulle
reproducera sig bara ett par månader senare. Dessa
koraller släppte sedan inte heller ut några äggceller
då de egentligen skulle göra det, trots att de
hade återhämtat sig från korallblekningen, och
i blekta koraller från andra arter var antalet
äggceller betydligt mindre än i individer som
inte hade blekts. Man har också visat att utvecklingen
av korallers könsorgan går långsammare vid de
temperaturer som orsakar korallblekning, samtidigt
som andra viktiga processer (exempelvis befruktning)
avbryts.
Alla dessa upptäckter pekar
på att korallblekning även resulterar i mer eller
mindre osynliga effekter som kanske inte uppfattas
direkt, och det kan spela en viktig roll i hur
väl korallrev återhämtar sig från massblekningar.
Förutom att koraller med reproduktiv potential
dör i samband med massblekningar kan alltså även
överlevande individer förlora möjligheten till
att reproducera sig, något som kan ha allvarliga
konsekvenser för hur många nya koraller som rekryteras.
I värsta fall kan drabbade korallarter helt förlora
möjligheten till att återuppbygga sina populationer
efter en period av massmortalitet. Detta var något
man oroade sig för i speciellt Indiska Oceanen
efter massblekningen 1997-98, men idag verkar
det alltså som om drabbade korallrev i vissa regioner
uppvisar en positiv återhämtning (se Massblekningen
1997-98).
Reducerad
produktivitet och kalkavlagring
Även om alla blekta koraller inte dör uppvisar
de istället en reducerad produktivitet, kalkavlagring
och förmåga till att reparera sig själva. Korallblekning
resulterar som sagt i en förlust av zooxantheller
hos drabbade koraller, och med dem försvinner
då också en viktig födokälla. Det resulterar i
en minskad fotosyntetisk produktivitet och tillväxt,
och eftersom koraller (tillsammans med zooxantheller)
är viktiga primärproducenter på ett korallrev
försvinner därmed också en stor del av korallrevets
produktivitet. Men förutom denna effekt påverkar
också en förlust av zooxantheller korallernas
kalkavlagring. Zooxanthellernas fotosyntes främjar
som bekant korallernas energimässigt kostsamma
kalkavlagring, och när den minskar i omfattning
i samband med korallblekning försvagas därmed
korallernas kalkskelett. Tillsammans med en reducerad
tillväxt kan det i sin tur resultera i en försämrad
konkurrenskraft hos blekta koraller, som då kan
få det svårare att konkurrera med exempelvis alger,
och korallrev drabbade av korallblekning kan på
så vis förlora sina koraller till förmån för alger.
I Karibien och östra Stilla Havet har också korallrev
som tidigare dominerades av koraller förvandlats
till korallrev som domineras av alger.
Interaktioner
mellan förhöjda vattentemperaturer och
andra hot
Den försämrade vitaliteten hos korallerna under
pågående korallblekning påverkar också deras moståndskraft
mot andra störningar. Sedimentation, utsläpp av
oorganiska näringsämnen, och andra störningar
som exempelvis destruktiva fiskemetoder kan i
kombination med förhöjda vattentemperaturer mycket
väl ta död på hela korallrev i vissa områden,
och i Karbien har man sett att bara en liten ökning
av vattentemperaturen förstör korallrev i kombination
med andra störningar. Utbrott av korallsjukdomar
(se Korallsjukdomar)
kan i vissa fall också kopplas till förhöjda vattentemperaturer,
och det är möjligt att larvutvecklingen hos den
effektiva korallpredatorn Acanthaster planci
främjas av ett varmare vatten. Det skulle i sin
tur kunna gynna utbrott av denna sjöstjärna (se
Sjöstjärnan
Acanthaster planci).
Förutom att andra hot kan intensifiera
effekterna av korallblekning kan de också påverka
återhämtningen från perioder av massblekning.
Exempelvis kan en försämrad vattenkvalitet påverka
befruktningsframgången hos koraller och därmed
begränsa korallernas och hela ekosystemets förmåga
att återhämta sig, och om tillväxten av alger
underlättas av förhöjda halter av oorganiska näringsämnen
kan rekryteringen av nya planula-larver försvåras.
Herbivora fiskar betar alger, men om populationerna
av dessa fiskar har minskat som ett resultat av
mänskliga aktiviteter kan det också påverka ett
korallrevs återhämtning från korallblekning och
massmortalitet. Det finns exempelvis fall då återhämtningen
har blivit mer utdragen eller till och med avstannat
i samband med en kraftig minskning av herbivorer,
och detta har, under en period på över ett decennium,
resulterat i en stadig övergång från korallrev
som tidigare dominerades av koraller till korallrev
som domineras av alger. För korallrev som hotas
av både överfiske och försämrad vattenkvalitet
kan återhämtningen därmed bli än mer besvärlig,
och det är därför viktigt att effektivt övervaka
mänskliga aktiviteter för att underlätta återhämtningen.
Fyra
möjliga framtida scenarier
En potentiell effekt relaterad till massblekningar
är att den biologiska mångfalden på korallrev
förändras. Lokalt utdöende av korallarter har
redan påträffats, men hur framtida förändringar
i artsammansättningen på korallrev kommer att
påverka korallrevens långsiktiga stabilitet är
dock oklart. Därför har fyra möjliga framtida
scenarier presenterats för att ge en klarhet i
hur en fortsatt global uppvärmning kommer att
påverka korallrev.
1. Koraller och andra
symbiotiska organismer kan acklimatisera sig till
både de förhöjda ytvattentemperaturerna och den
sänkta aragonitmättnaden i havet (se Övriga
hot relaterade till globala klimatförändringar).
Detta scenario skulle alltså resultera i att korallerna
överlever och att någon ekosystemsförändring inte
inträffar. Eftersom koraller runt om i världen
utsätts för en allt intensivare korallblekning
skulle dock en sådan acklimatisering vara omöjlig,
och korallerna verkar inte heller acklimatisera
sig tillräckligt snabbt (se Kan
inte korallerna anpassa sig?).
2. Den ökande dödligheten
bland korallerna resulterar i att korallernas
genomsnittliga livslängd förkortas. Inga korallarter
försvinner, men individerna blir istället yngre.
3. Massiva och långsamt
växande koraller, som är mer motståndskraftiga
mot korallblekning, ersätter de snabbväxande förgrenade
korallerna. Detta är ett scenario som troligtvis
redan har iscensatts, och att vissa korallarter
börjar bli mer och mer ovanliga understryker det
bara ytterligare.
4. Korallerna ersätts
av andra organismer som alger. Detta har redan
rapporterats från både Karibien och östra Stilla
Havet, och troligtvis skulle dessa förändringar
av ekosystemet drabba korallrev i alla havsområden
inom bara ett par hundra år så länge vattentemperaturen
fortsätter att stiga och korallerna är oförmögna
att anpassa sig till de ny förhållandena. När
dessa ekosystemsförändringar väl har genomförts
kan det ta flera hundra år innan de drabbade korallreven
domineras av koraller igen.
Konsekvenserna
av korallblekning för andra organismer
Koraller (tillsammans med sina endosymbionter)
står som sagt för en stor del av primärproduktionen
på korallrev. Zooxanthellerna absorberar solljus
och för den energin, genom fotosyntes, vidare
till organiska föreningar som korallerna tar upp,
och korallerna blir sedan i sin tur föda åt en
mängd olika organismer. Förutom detta bygger korallerna
också upp en komplex struktur som förser majoriteten
av alla korallrevsorganismer med skydd. Följaktligen
skulle alltså en reduktion av mängden koraller
och korallarter i samband med korallblekning resultera
i stora konsekvenser för de flesta organismer
som associeras med korallrev.
Det är med andra ord tydligt
att den globala uppvärmningen kommer att ha märkbara
effekter på korallrevets biodiversitet, men lite
är fortfarande känt om de mer detaljerade effekterna,
och studier som har tittat på hur korallrevsfiskar
reagerar på korallblekning har visat på
varierande resultat. I en undersökning från Seychellerna
såg man exempelvis att vissa fiskarter ökade i
antal efter massblekningen 1997-98 medan en annan
studie visade att ett kraftigt minskat antal koraller,
delvis som ett resultat korallblekning, resulterade
i att majoriteten av alla fiskarter minskade i
antal. Men trots dessa resultat är det dock uppenbart
att de korallrevsorganismer som på något sätt
är direkt beroende av koraller, exempelvis genom
att ha dem som sin enda födokälla, kommer att
drabbas hårdare än andra. Oxymonacanthus longirostris,
en fisk som endast äter Acropora-polyper,
försvann exempelvis snabbt från korallreven runt
Okinawa efter massblekningen 1997-98. Att vissa
korallarter är mer mottagliga för korallblekning
kan dessutom få ytterligare konsekvenser. Förgrenade
koraller, som har sämst motståndskraft, förser
flera arter, inklusive ett stort antal fiskar
och ryggradslösa djur, med en viktig livsmiljö,
och om de då försvinner kan det påverka även dessa
organismer. Som tidigare nämnts finns det dock
också andra faktorer som påverkar den framtida
artsammansättningen av koraller, och även massiva
korallers förmåga att återhämta sig kan komma
att försämras om massblekningar blir mer vanligt
i framtiden. Den allmänna uppfattning idag är
emellertid att förgrenade koraller kommer att
drabbas hårdast av den globala uppvärmningen med
förändringar i artsammansättningen som resultat
(se scenario 3 under Fyra
möjliga framtida scenarier), och att vi troligtvis
kommer att se en omfattande degradering av korallrev
i framtiden.
Troligtvis kommer också människan
att drabbas eftersom en lägre produktivitet i
kombination med förluster av kritiska livsmiljöer
för flera korallrevsorganismer kan resultera i
reducerade fiskfångster. Det beror på att de fiskar
som är beroende av koraller också är föda åt de
större fiskarter som fiskerier fångar, och korallblekning
skulle därför ha en indirekt effekt på speciellt
småskalig fiskenäring. Men även om det finns exempel
på hur massblekningen 1997-98 hade en negativ
påverkan på olika fiskpopulationer, har påverkan
på fiskfångster och inkomster varit mycket svårare
att dokumentera. Fångsterna i tropiska vatten
håller i och för sig på att minska, men det beror
snarare på det överfiske man ser idag än på förhöjda
vattentemperaturer. Det finns dock en påtaglig
risk för att detta problem skulle förvärras om
den globala uppvärmningen fortsätter och massblekningar
blir allt vanligare och intensiva, och då skulle
miljontals människor i speciellt tropiska utvecklingsländer
troligtvis förlora sin viktigaste proteinkälla.
Korallblekning kan också ha
en inverkan på turismen då skadade korallrev inte
har samma dragningskraft som friska. Under åren
1998-2001, direkt efter massblekningen 1997-98,
uppskattades exempelvis de ekonomiska förlusterna
för Seychellerna i uteblivna turister till 9,7
miljoner amerikanska dollar, och enligt en studie
från Australien kan skador orsakade av korallblekning
resultera i att turistindustrin förlorar miljardbelopp
fram till år 2020.
Även fåglar, reptiler och marina
däggdjur förväntas drabbas om produktiviteten
på korallrev minskar, och man har redan sett fall
av kraftig mortalitet bland sjöfåglar i samband
med ENSO-år.
Kan
inte korallerna anpassa sig?
En kritisk del av korallernas framtida öde beror
på hur de, och deras zooxantheller, kommer att
reagera på en stigande havsvattentemperatur. Generellt
sett kan marina organismer reagera på två olika
sätt vid temperaturförändringar. För det första
kan individuella organismer acklimatisera sig
till temperaturförändringar genom att modifiera
sina cellulära processer så att de fortsätter
att fungera även vid de nya temperaturerna. Exempelvis
skulle koraller kunna ändra sin fysiologi (eller
biokemi) så att de kan tolerera högre vattentemperaturer.
Det andra sättet som organismer kan reagera på
är att selektera de individer i en population
som tål de nya temperaturerna bäst. Detta skulle
alltså innebära att de mest tåliga individerna
skulle överleva medan de andra, mindre tåliga
skulle dö eller misslyckas med sin reproduktion.
På det sättet skulle nya så kallade adaptioner
evolveras fram med tiden och i fallet med korallerna
och zooxanthellerna skulle de då på sikt kunna
klara av högre vattentemperaturer.
På frågan om koraller och deras
zooxantheller kan acklimatisera och/eller adaptera
sig till förändrade havsvattentemperaturer beror
svaret på hur lång tid det skulle ta för dem att
göra de nödvändiga förändringarna. Växter och
djur kan acklimatisera sig till temperaturförändringar
på bara några timmar eller dagar, oberoende av
generationstiden. Det kan dock ta flera hundra
eller tusen år för samma organismer att adaptera
sig till samma temperaturförändringar, och i det
här fallet beror också adaptionstiden på organismernas
generationstid. Koraller och fiskar, som har en
lång generationstid, skulle således behöva flera
tusen år på sig innan de har evolverat fram nya
adaptioner medan exempelvis kortlivade bakterier
bara skulle behöva några dagar. Man har också
visat att organismer med kort generationstid överlever
större miljöförändringar bättre än organismer
med lång generationstid.
Koraller och zooxantheller i
olika geografiska områden har under tusentals
år genetiskt adapterat sig till olika vattentemperaturer.
Koraller från Eniwetokatollen, där den genomsnittliga
vattentemperaturen är 28,5°C, överlevde exempelvis
en exponering för 35,6-gradigt vatten under 10
timmar, medan de flesta koraller från Hawaii,
där den genomsnittliga vattentemperaturen ligger
på 24,5°C, inte överlevde en vattentemperatur
på 32,4°C. Koraller från kallare områden bleknar
alltså vid lägre vattentemperaturer än vad koraller
från varma områden gör. Denna observation att
koraller är anpassade till lokala temperaturförhållanden
är emellertid ett universellt särdrag som gäller
för alla organismer, men skulle gener (larver)
från koraller i varmt vatten transporteras med
havsströmmarna till områden med kallare vatten,
kan motståndskraften mot korallblekning öka i
dessa områden. Det är dock inte troligt att dessa
förändringar på populationsnivå kommer att inträffa
snabbt, men det finns ett ökande antal indikatorer
på att koraller nu faktiskt rör sig mot högre
latituder, samtidigt som kvarvarande individer
dör i det allt varmare vattnet i tropikerna. Att
arealen av grunda bottnar och koncentrationen
av karbonatjoner minskar, samtidigt som ljusintensiteten
avtar, från ekvatorn och upp mot högre latituder
innebär emellertid att detta inte är lösningen
för världens koraller och korallrev.
Det finns också obervationer
på att vissa individer i en population av koraller
är mer värmetåliga än andra, och det skulle kunna
innebära att populationer på egen hand kan bli
mer värmetåliga om dessa individer selekteras.
Koraller på ett korallrev är dock ofta genetiskt
kopplade till varandra, vilket antyder att den
genetiska diversiteten inte alltid är tillräckligt
hög på vissa korallrev för att de mest värmetåliga
individerna i en population skall kunna förse
populationen med den genetiska bas som kommer
att behövas i ett allt varmare havsvatten. Eftersom
vissa korallrev också har drabbats av alla globala
massblekningar sedan 1979 verkar inte heller koraller
och zooxantheller förändra sin genetiska uppsättning
till en mer värmetålig version tillräckligt snabbt.
"Adaptove
Bleaching Hypothesis" och acklimatisering
I början av 1990-talet kom den så kallade "Adaptive
Bleaching Hypothesis" som menade att koraller
kanske kan byta ut sina zooxantheller mot mer
värmetåliga typer, och att det då skulle resultera
i en ökad överlevnad i samband med temperaturstress.
Värmestressade koraller skulle då aktivt stöta
ut den redan existerande zooxanthelltypen för
att istället börja utnyttja en mer värmetålig
typ medan värmestressen fortfarande finns närvarande.
Detta skulle då förklara förlusterna av zooxantheller
i samband med korallblekning. Det finns dock inga
obervationer på att koraller faktiskt aktivt stöter
ut sina zooxantheller och istället börjar utnyttja
nya under pågående värmestress, och det är därför
mer troligt att förlusterna av zooxantheller snarare
är relaterad till utstötningen av korallpolypernas
egna celler (se De
biokemiska processerna bakom blekningen).
Kritiker av denna hypotes har också argumenterat
för att den massmortalitet man ibland ser i samband
med korallblekning inte kan anses vara ett adaptivt
beteende. Koraller kan dock fortfarande byta ut
sina zooxantheller mot en mer värmetålig typ efter
det att korallblekningen har ägt rum och i samband
med att deras populationer av zooxantheller skall
återuppbyggas, och under de första åren av 2000-talet
kom det bevis på att vissa koraller faktiskt kan
acklimatisera sig genom att byta ut de zooxantheller
som dominerar för tillfället (typ C) till en annan,
mer värmetålig variant (typ D). Detta har oberverats
på korallrev i både Karibien och Stilla Havet,
och med tanke på denna spridning kan korallrev
faktiskt ha en större möjlighet att klara av framtida
klimatförändringar än vad som tidigare var trott.
Exempelvis visade en studie från Stora Barriärrevet
att den vanliga korallarten Acropora millepora
kunde tåla temperaturer på 1-1,5°C över dess normala
temperaturmarginal när den bytte symbionter från
zooxantheller av typ C till zooxantheller av typ
D, och områden som inte drabbades lika hårt som
andra havsområden under massblekningen 1997-98
inneslöt också koraller som hade mer zooxantheller
av typ D än typ C. Det är dock fortfarande inte
känt om korallerna tar upp de nya zooxanthellerna
från det omgivande vattnet eller om de små populationer
av den mer värmetåliga zooxanthelltypen som redan
återfinns inne i korallernas vävnad helt enkelt
får öka i antal. Tyvärr har dessa positiva resultat
fått dämpas av en mer aktuell studie från 2006.
Den visade att alla korallarter inte har möjligheten
att öka mängderna av den mer värmetåliga typ D.
För att kunna göra detta krävs det nämligen att
korallen kan ingå i ett symbiotiskt förhållande
med mer än en typ, och av de 442 korallarter som
ingick i studien hade endast 23 procent denna
egenskap. Huruvida väl koraller kan anpassa sig
på detta vis återstår därför att se.
Koraller skulle dock kunna förändra
sin fysiologi (eller biokemi) efter den globala
uppvärmningen och acklimatisera sig på detta vis.
Dessa förändringar skulle då ta allt från timmar
till dagar att genomföra och vara antingen tillfälliga
eller permanenta. En korall skulle på så vis tåla
även framtida stress, exempelvis höga vattentemperaturer
under nästkommande sommar. Det finns också exempel
på hur koraller i kustnära områden acklimatiserar
sig till en mer stressfull miljö, och korallpolyper
som har utsatts för höga ljusintensiteter har
också visat sig vara mer resistenta mot korallblekning
i samband med förhöjda ytvattentemperaturer, vilket
antyder att koraller kan acklimatisera sig till
höga ljusintensiteter. Att acklimatisera sig innebär
dock en energimässig kostnad för korallen, något
som kan drabba andra processer, exempelvis reproduktionen,
och säsongsvariationer och störningar i samband
med ENSO-år kan försvåra acklimatiseringen. En
fysiologisk förändring behöver som sagt var inte
heller vara permanent, och eftersom perioder av
varmt vatten vanligtvis åtföljs av perioder med
kallt vatten hinner de koraller som har acklimatiserat
sig till varmare vatten förlora dessa nya egenskaper
under den påföljande kalla perioden, och de kan
lättare drabbas av korallblekning när det återigen
blir varmt. Dessutom måste acklimatiseringen följa
den stigande vattentemperaturen, vilket ter sig
allt mer otroligt i och med de framtida ytvattentemperaturer
som förutses, och vissa korallrev har som sagt
drabbats av alla globala massblekningar sedan
1979, vilket tyder på att ingen omfattande acklimatisering
har skett under de senaste decennierna.
En annan form av acklimatisering
kan vara utnyttjandet av de fluorescerande pigment
som återfinns i flera korallers vävnad. Koraller
skulle kunna förändra tätheten eller placeringen
av dessa pigment inne i deras vävnad för att på
så vis reducera skadan på zooxanthellernas fotosyntes
i samband med värmestress och minska risken för
korallblekning. Pigmenten skulle då helt enkelt
utnyttjas till att skugga zooxanthellerna snarare
än att förbättra tillgängligheten
av ljus, vilket är en annan viktig funktion
hos pigmenten (se Korallers
biologi). Att vissa koraller, som verkar vara
mer resistenta än andra mot blekning, uppvisar
intensiva pastellfärger i samband med korallblekning
tyder också på att denna pigmentering spelar en
roll i att hjälpa koraller acklimatisera sig till
värmestress. Forskningen kring fluorescerande
pigment är ett aktivt fält och det återstår
att se vilken långsiktig betydelse de har
för korallers fortsatta välstånd.
Sammanfattningsvis finns det
alltså idag inga övertygande bevis på någon storskalig
förändring av korallers och zooxanthellers tolerans
mot förhöjda vattentemperaturer eller att någon
form av anpassning mot korallblekning förekommer,
även om det finns en varierande optimism inom
forskarvärlden vad gäller korallers och korallrevs
möjligheter att anpassa sig till de stigande vattentemperaturerna.
Övriga
hot relaterade till globala klimatförändringar
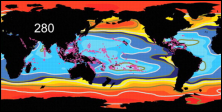
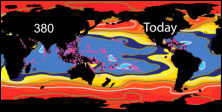
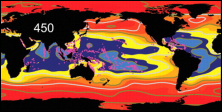
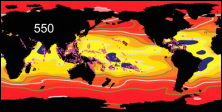
Koldioxidhalten ökar kraftigt i atmosfären (till
och med kraftigare än vad som tidigare har förväntats),
och ungefär en fjärdedel av den koldioxid mänskliga
aktiviteter släpper ut absorberas av havet. En
förhöjd halt koldioxid i havet påverkar aragonitmättnaden
och surheten (se Inblick:
Mer om koldioxid, surhet och sänkt aragonitmättnad
nedan), och det har på sistone visat sig vara
av stor betydelse för korallrevens fortsatta välstånd.
Genom att utnyttja siffror över förväntade framtida
koldioxidhalter har man i två studier från 1999
visat att aragonitmättnaden kommer att sjunka
med 30 procent i världens tropiska hav, och eftersom
mineralen aragonit (en av två former av kalciumkarbonat,
där den andra är kalcit) är en viktig beståndsdel
i både korallers och andra organismers kalkskelett,
tror man att hastigheten på korallers och andra
organismers kalkavlagring i tropiska vatten därför
kommer att sjunka med 14 till 30 procent till
år 2050. Även senare studier från början av 2000-talet
har uppvisat liknande resultat; skulle den förindustriella
koldioxidhalten fördubblas, vilket förväntas till
2050, kommer korallers kalkavlagring att reduceras
med 10-40 procent. Detta skulle då resultera i
ett försvagat kalkskelett och en försämrad tillväxt
bland korallerna. I en annan studie från 2004
visade man emellertid att effekten av en sänkt
aragonitmättnad kan mer än kompenseras av en stigande
havsvattentemperatur. Koraller växer snabbare
i varmare vatten, och författarnas förutsägelse
var därför att koraller kommer att växa 35 procent
snabbare år 2100. Studien beaktade dock inte möjligheten
att koraller i varmare vatten bleks, samtidigt
som den utfördes på endast en enda korallart som
dessutom var den mest värmetåliga. Författarna
har därför fått kritik för att de inte har tagit
hänsyn till viktiga faktorer och utelämnat andra,
mindre värmetåliga korallarter. Däremot skulle
förhöjda havsvattentemperaturer kunna hämma världshavens
koldioxidupptag. Lösligheten av koldioxid i vatten
är nämligen omvänt relaterad till vattnets temperatur.
Koldioxid löser sig därför inte lika väl i varmt
vatten som i kallt, vilket innebär att de effekter
en förhöjd halt koldioxid i atmosfären har på
korallrev kan reduceras om temperaturen stiger
i havet.
Den generella uppfattningen
är dock att en sänkt aragonitmättnad och förhöjda
ytvattentemperaturer kommer att ha negativa effekter
på tillväxten hos världens korallrev. Ett korallrevs
tillväxt styrs av en balans mellan den totala
kalkavlagringen och fysisk och biologisk erosion,
så en minskad kalkavlagring skulle då troligtvis
resultera i att erosionen blir större än kalkavlagringen,
och att korallrevet skulle minska i storlek. Detta
är i så fall något som kommer att inträffa i flera
havsområden i framtiden (se Inblick:
Mer om koldioxid, surhet och sänkt aragonitmättnad
nedan) med konsekvenser för både människan och
andra marina ekosystem; drabbade korallrev skulle
inte längre fungera lika väl som kustskydd, och
andra marina ekosystem (sjögräsbäddar och mangroveskogar)
som förlitar sig på korallrev skulle påverkas
negativt. Dessutom skulle en ökad erosion kunna
påverka ett korallrevs återhämtning från massblekningar
genom att skapa ett mer instabilt underlag för
nya koraller att kolonisera.
En annan konsekvens av den globala
uppvärmningen är att havsvattennivån stiger. Enligt
en uppskattning från 1998 har havsvattennivån
stigit med 10-25 centimeter under de senaste 100
åren, och enligt samma studie kommer den att fortsätta
att stiga med ytterligare 15-95 centimeter under
det närmaste seklet; senare studier av IPCC (FN:s
klimatorgan) från 2007 har bekräftat att den globala
havsvattennivån steg med i genomsnitt 17 centimeter
under 1900-talet, och att den maximala höjningen
av havsvattennivån fram till år 2100 är 51 centimeter,
baserat på ett av deras scenarier. Koraller trivs
som bekant bäst i grunda vatten där deras zooxantheller
kan ta till sig solljus och fotosyntetisera. En
förhöjd havsvattennivå skulle således resultera
i att koraller i djupare vatten får det svårare
att tillväxa, och till slut skulle de inte längre
kunna hålla sig vid liv på grund av den låga solinstrålningen.
Samtidigt frigörs dock mer yta för koralltillväxt
på de ytligaste delarna av ett korallrev, och
det är därför möjligt att dessa delar av korallrevet
då istället gynnas av en stigande havsvattennivå.
Eftersom föroreningar och sediment kan sköljas
ner från närliggande kustområden i samband med
att havsvattennivån stiger, kan emellertid en
förhöjd havsvattennivå också innebära att andra
störningar skapas.
Det är emellertid svårt att
förutse om en förhöjd havsvattennivå verkligen
kan dränka koraller och korallrev eftersom tillväxten
mellan både olika korallarter och koraller och
korallrev är så varierande. Snabbväxande förgrenade
koraller, som har en vertikal tillväxt på uppemot
10 centimeter på ett år, kommer inte att ha några
problem med att klara av dessa förändringar, men
problemet blir mer tydligt då man tar hänsyn till
mer massiva och långsamt växande koraller, och
till hela korallrev. Skulle havsvattenivån stiga
med maximala 0,95 centimeter per år (i enlighet
med studien från 1998) matchar det långsamt växande
korallers tillväxt som ligger på uppskattningsvis
en centimeter per år. Om dessa koraller dessutom
skulle få sin tillväxt reducerad av korallblekning
och andra störningar, kan förändringarna i havsvattennivån
bli större än korallernas tillväxt. Tillväxten
hos hela korallrev är dock oftast betydligt långsammare
än hos enskilda koraller, och om både erosionen
och förekomsten och intensiteten av mänskliga
aktiviteter och massblekningar ökar i framtiden,
kan tillväxten avstanna helt eller till och med
bli negativ. Då krävs inga stora förändringar
i havsvattennivån innan vi börjar se effekter
på drabbade korallrev, och eftersom även de mest
konservativa scenarierna över framtida havsvattenivåer
pekar på att havsvattennivån kommer att stiga
globalt, kan alltså en kombination av utsläpp
av växthusgaser och andra mänsliga aktiviteter
på sikt rent av hota korallrevens hela existens.
|
|
 Koraller
och korallrev Koraller
och korallrev |
 |
|
|
|
 Globala
klimatförändringar och Globala
klimatförändringar och
korallblekning |
 |
|
|
|